
Molecular Modeling: F1F0-ATPSynthase
 Die F1F0-ATPSynthase ist ein molekularer Motor, welcher in Bakterien, Pflanzen und Tieren vorkommt und an bedeutenden Energieumwandlungsprozessen mitwirkt. In der bakteriellen Plasmamembran, der Thylakoidmembran der Chloroplasten und der inneren Mitochondrienmembran gelegen, wandelt die F1F0-ATPSynthase einen elektrochemischen Protonengradienten in chemische Energie "in Form von ATP" um. Der dafür benötigte Protonengradient stammt bei Pflanzen und photosynthetisch aktiven Bakterien aus der Lichtreaktion der Photosynthese, bei Tieren aus der Atmungskette.
Die F1F0-ATPSynthase ist ein molekularer Motor, welcher in Bakterien, Pflanzen und Tieren vorkommt und an bedeutenden Energieumwandlungsprozessen mitwirkt. In der bakteriellen Plasmamembran, der Thylakoidmembran der Chloroplasten und der inneren Mitochondrienmembran gelegen, wandelt die F1F0-ATPSynthase einen elektrochemischen Protonengradienten in chemische Energie "in Form von ATP" um. Der dafür benötigte Protonengradient stammt bei Pflanzen und photosynthetisch aktiven Bakterien aus der Lichtreaktion der Photosynthese, bei Tieren aus der Atmungskette.Die Bezeichnungen F1 und F0 stehen jeweils für die Bereiche der ATPSynthase, welche sich außerhalb bzw. innerhalb der Membran befinden. Es wurden bereits die Strukturen von Einzelteilen der F1F0-ATPSynthasen verschiedener Organismen augeklärt. Durch das Zusammenlegen 4 einzelner Strukturen aus E. coli und B. taurus (Rind) ist es möglich, sich ein aufschlussreiches Bild zu machen von der Wirkungsweise des allgegenwärtigen molekularen Motors.
VMD-Darstellung herunterladen mit integrierten Animationen und interaktiven Buttons zum schnellen Wechsel zwischen verschiedenen Darstellungen)!
 Struktur
Struktur
Wie ein makroskopischer Motor besteht  auch die F1F0-ATPSynthase aus einem Stator und einem Rotor, welche sich bei Betrieb relativ zueinander bewegen. Der Stator besteht aus einem kleinen transmembranalen "Verankerungsanteil", einem großen kugelförmigen Teil, welcher die drei aktiven Zentren beinhaltet und aus der Membran herausragt, und einem langen "Verbindungsstiel" zwischen den beiden Teilen. Den Stator sollte man sich vorstellen als die eher unbewegliche Hälfte, welche den Rotor festhält. Der Rotor ist ein annähernd rotationssymmetrisches Gebilde, welches im Stator verankert ist und während seiner Rotation die notwendige Energie zur ATP-Synthese auf diesen überträgt:
auch die F1F0-ATPSynthase aus einem Stator und einem Rotor, welche sich bei Betrieb relativ zueinander bewegen. Der Stator besteht aus einem kleinen transmembranalen "Verankerungsanteil", einem großen kugelförmigen Teil, welcher die drei aktiven Zentren beinhaltet und aus der Membran herausragt, und einem langen "Verbindungsstiel" zwischen den beiden Teilen. Den Stator sollte man sich vorstellen als die eher unbewegliche Hälfte, welche den Rotor festhält. Der Rotor ist ein annähernd rotationssymmetrisches Gebilde, welches im Stator verankert ist und während seiner Rotation die notwendige Energie zur ATP-Synthese auf diesen überträgt:
auch die F1F0-ATPSynthase aus einem Stator und einem Rotor, welche sich bei Betrieb relativ zueinander bewegen. Der Stator besteht aus einem kleinen transmembranalen "Verankerungsanteil", einem großen kugelförmigen Teil, welcher die drei aktiven Zentren beinhaltet und aus der Membran herausragt, und einem langen "Verbindungsstiel" zwischen den beiden Teilen. Den Stator sollte man sich vorstellen als die eher unbewegliche Hälfte, welche den Rotor festhält. Der Rotor ist ein annähernd rotationssymmetrisches Gebilde, welches im Stator verankert ist und während seiner Rotation die notwendige Energie zur ATP-Synthese auf diesen überträgt: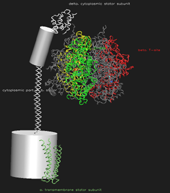{kind=link}
{kind=link}
Katalysevorgang
Der Stator der F1F0-ATPSynthase enthält 3 aktive Zentren, an denen ATP aus ADP und anorganischem Phosphat unter Abspaltung von Wasser gebildet wird. Die auf diese Weise entstehende Anhydridbindung speichert die freie Energie, welche zum Antrieb verschiedener Reaktionen verwendet werden kann.
Die aktiven Zentren befinden sich in dem kugelförmigen Teil des Stators, welcher bei Bakterien in das Zellinnere und bei Mitochondrien von Pflanzen und Tieren in die Matrix (innen) ragt, und bei der Thylakoidmembran der Chloroplasten (duch die Entstehung der Thylakoidmembran bedingt) auf der Außenseite liegt; der Ausdruck "cytoplasmatisch" bezieht sich für diesen Teil auf die Lage im Bakterium (ins Zellinnere gerichtet). Dieser Teil ist aus jeweils drei Kopien zwei verwandter Untereinheiten aufgebaut (3 mal α, 3 mal β), welche wie die einzelnen Stücke einer Zitrusfrucht angeordnet sind und sich in ihrer Reihenfolge abwechseln (αβαβαβ). Während die 3 aktiven Zentren an der Grenzfläche jeder zweiten Untereinheit sitzen, sind lediglich die β-Untereinheiten katalytisch aktiv.
Wird vom Rotor eine mechanische Kraft übertragen, so werden in einem durch Konformationsänderungen bewirkten Zyklus zuerst ADP und anorganisches Phosphat in einer Tasche am aktiven Zentrum gebunden, die Reaktion zu ATP durch Schließen der Tasche und Ausschluss von Wassermolekülen aus der Tasche begünstigt und zuletzt das Reaktionsprodukt (ATP) aus der wieder geöffneten Tasche freigesetzt, um wieder neue Edukte (ADP und anorganisches Phosphat) aufzunehmen. Diese drei Schritte des Reaktionszyklus entsprechen drei Zuständen der Bindetasche, welche der Reihe nach durchlaufen werden: L(oose) zur Bindung der Edukte, T(ight) zur Bildung von ATP und O(pen) zur Freisetzung von ATP.
Diese drei Schritte des Reaktionszyklus entsprechen drei Zuständen der Bindetasche, welche der Reihe nach durchlaufen werden: L(oose) zur Bindung der Edukte, T(ight) zur Bildung von ATP und O(pen) zur Freisetzung von ATP.Nach dem akzeptierten Binding-Change-Mechanismus befindet sich immer alle drei aktiven Zentren in einem anderen der drei Zustände, in einer F1F0-ATPSynthase ist also immer jeweils ein aktives Zentrum in der L-,T- und O-Konformation vorhanden. Die dazugehörigen β-Unterheiten sind in der Abbildung farbig hervorgehoben (L:gelb, T:rot, O:grün).
{kind=link}
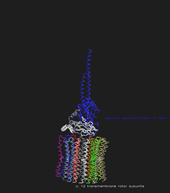
Die Kraftübertragung des Rotors auf die α- und β-Untereinheiten des Stators erfolgt üner die stabförmige γ-Untereinheit, welche in einer Hohlröhre im Zentrum des kugelförmigen Teils des Stators liegt. Durch die Assymmetrie der γ-Untereinheit können die drei verschiedenen aktiven Zentren zu einem Zeitpunkt in den drei verschiedenen Konformationen (L,T,O) vorliegen. Die Hohlröhre steht mit der γ-Untereinheit nur über schwache hydophobe Wechselwirkungen in Kontakt, sodass die Reibung bei der Rotation minimiert wird und ein unglaublich hoher Wirkungsgrad bei der Energieumwandlung resultiert.
Angetrieben wird der Rotor durch den symmetrischen membranalen Teil, einem radörmigen Gebilde aus 10-12 identischen c-Untereinheiten. Diese stehen an einer Seite mit der membranalen a-Untereinheit des Rotors in Kontakt. An dieser Stelle wird das Rad aus c-Untereinheiten angetrieben: Auf einem noch nicht ganz bekannten Weg werden Protonen zu der Kontaktstelle der a-Untereinheit und einer c-Untereinheit geschleust. Dort befindet sich eine positiv geladene Aminosäure (Arg, a-Untereinheit) und eine negativ geladene Aminosäure (Asp, c-Untereinheit), welche sich aufgrund ihrer gegensätzlichen Ladung anziehen und das Drehen des Rotors verhindern.
Durch die Bindung des Protons an das negativ geladene Asp wird dessen Ladung neutralisiert und die elektostatische Bindung wird aufgelöst.  Durch eine Konfomationsveränderung schwingt die c-Untereinheit an der a-Untereinheit vorbei, ein Zurückschwingen ist wegen eines Ratschen-Mechanismus nicht ohne Weiteres möglich. Die dabei entstehende Kraft wird auf den gesamten Rotor übertragen und bewirkt ein kleine Rotation. Der a-Untereinheit liegt nun die nächste c-Untereinheit gegenüber. Liegen genau 12 c-Untereinheiten vor, so muss dieser elementare Schritt genau zwölfmal stattfinden, damit der Rotor eine gesamte Drehung vollbringt. Währenddessen durchläuft jedes aktive Zentrum im Stator alle 3 Konformationen und es werden 3 Moleküle ATP synthetisiert:
Durch eine Konfomationsveränderung schwingt die c-Untereinheit an der a-Untereinheit vorbei, ein Zurückschwingen ist wegen eines Ratschen-Mechanismus nicht ohne Weiteres möglich. Die dabei entstehende Kraft wird auf den gesamten Rotor übertragen und bewirkt ein kleine Rotation. Der a-Untereinheit liegt nun die nächste c-Untereinheit gegenüber. Liegen genau 12 c-Untereinheiten vor, so muss dieser elementare Schritt genau zwölfmal stattfinden, damit der Rotor eine gesamte Drehung vollbringt. Währenddessen durchläuft jedes aktive Zentrum im Stator alle 3 Konformationen und es werden 3 Moleküle ATP synthetisiert:
Durch eine Konfomationsveränderung schwingt die c-Untereinheit an der a-Untereinheit vorbei, ein Zurückschwingen ist wegen eines Ratschen-Mechanismus nicht ohne Weiteres möglich. Die dabei entstehende Kraft wird auf den gesamten Rotor übertragen und bewirkt ein kleine Rotation. Der a-Untereinheit liegt nun die nächste c-Untereinheit gegenüber. Liegen genau 12 c-Untereinheiten vor, so muss dieser elementare Schritt genau zwölfmal stattfinden, damit der Rotor eine gesamte Drehung vollbringt. Währenddessen durchläuft jedes aktive Zentrum im Stator alle 3 Konformationen und es werden 3 Moleküle ATP synthetisiert:- ATP-Synthase: Stillstand
- ATP-Synthase: Rotation
- ATP-Synthase: Elementarschritt zur Erzeugung einer Rotation
{kind=link}
Nach einer vollen Rotation wird das von der c-Untereiheit des Rotors aufgenommene Proton auf der anderen Seite der Membran abgegeben. Damit der gesamte Vorgang in der beschriebenen Richtung abläuft, muss auf dieser Seite die Protonenkonzentration geringer sein als auf der Seite der Protonenaufnahme. Durch den Fluss von Protonen über die F1F0-ATPSynthase wird somit der Protonengradient über die entsprechende Membran abgebaut und die in diesem Gradienten gespeicherte Energie in Form von chemischer Energie in der energiereichen Verbindung ATP gespeichert (Chemiosmotische Hypothese).
Neben den Motor-Eigenschaften der F1F0-ATPSynthase tragen auch Nicht-Motor-Eigenschaften zur Optimierung der Energieumwandlung bei.
Die verwendeten Strukturen stammen von der Protein Datenbank (RCSB PDB):
H.M.Berman, J.Westbrook, Z.Feng, G.Gilliland, T.N.Bhat, H.Weissig, I.N.Shindyalov, P.E.Bourne
The Protein Data Bank
Nucleic Acids Research, 28 pp. 235-242 (2000)
H.M.Berman, J.Westbrook, Z.Feng, G.Gilliland, T.N.Bhat, H.Weissig, I.N.Shindyalov, P.E.Bourne
The Protein Data Bank
Nucleic Acids Research, 28 pp. 235-242 (2000)
- PDB ID: 1C17
- Rastogi, V.K., Girvin, M.E.
- Structural changes linked to proton translocation by subunit c of the ATP synthase.
- Nature 402 pp.263-268 (1999)
- PDB ID: 1E79
- Gibbons, C., Montgomery, M.G., Leslie, A.G., Walker, J.E.
- The structure of the central stalk in bovine F(1)-ATPase at 2.4 A resolution.
- Nat.Struct.Biol. 7 pp.1055-1061 (2000)
- PDB ID: 1L2P
- Del Rizzo, P.A., Bi, Y., Dunn, S.D., Shilton, B.H.
- The "second stalk" of Escherichia coli ATP synthase: structure of the isolated dimerization domain.
- Biochemistry 41 pp.6875-6884 (2002)
- PDB ID: 2A7U
- Wilkens, S., Borchardt, D., Weber, J., Senior, A.E.
- Structural Characterization of the Interaction of the delta and alpha Subunits of the Escherichia coli F(1)F(0)-ATP Synthase by NMR Spectroscopy
- Biochemistry 44 pp.11786-11794 (2005)